It's been a few years since I wrote a post about alvarezsaurids on this blog, in which I discussed the interpretation of these dinosaurs as specialized insectivores that used their unusual forelimbs to dig for social insects, and why I happened to think that that was the most convincing model for their lifestyle and ecology. Back then, I had little expectation that there would be much alvarezsaurid news in the immediate future, so it was a pleasant surprise to see an alvarezsaurid featured prominently and portrayed as a termite-eating specialist in the high-profile documentary series Prehistoric Planet last year. Not only that, but many studies on alvarezsaurids have been published in the intervening time. Has this newfound information updated and modified my thoughts on how alvarezsaurids lived? Let's take a look.
The New Alvarezsaurs on the Block
To start off, several new alvarezsaurid taxa were named in recent years, including Trierarchuncus from the Hell Creek Formation of the western United States (Fowler et al., 2020; Freimuth and Wilson, 2021), Dzharaonyx from the Bissekty Formation of Uzbekistan (Averianov and Sues, 2021), and Khulsanurus and Ondogurvel from the Barun Goyot Formation of Mongolia (Averianov and Lopatin, 2022a; Averianov and Lopatin, 2022b). The braincase of an indeterminate alvarezsaurid from the Qiupa Formation of China was also described (Agnolín et al., 2022), and the anatomy of Parvicursor was reevaluated in detail (Averianov and Lopatin, 2021).
 |
| Nearly complete thumb claw of Trierarchuncus, from Fowler et al. (2020). |
In general, most of these new findings don't substantially change what I previously talked about, but it's great to have more alvarezsaur anatomy to work with. Trierarchuncus deserves mention, however, in that one of the specimens referred to it represents the most completely preserved alvarezsaurid thumb claw to date. I noted in my previous post on alvarezsaurs that complete alvarezsaurid thumb claws are hard to come by, making it difficult to compare their shape to the claws of modern animals. This Trierarchuncus specimen reveals that at least some alvarezsaurid thumb claws were much more sharply curved than formerly expected, in line with their hypothesized function in hook-and-pull digging.
 |
| Reconstructed curvature of the thumb claw of Mononykus based on that of Trierarchuncus, from Fowler et al. (2020). |
In addition, comparing the claws of differently sized Trierarchuncus individuals suggests that as alvarezsaurids aged, the bony core of their thumb claws became wider and gained a roughened texture (possibly induced by stress) near where the claw attached to the rest of the thumb, consistent with the use of the claws in strenuous activity (Fowler et al., 2020; Freimuth and Wilson, 2021).
Turning Tail
As I mentioned in my previous post, a long tail for balance can be a helpful adaptation to a hook-and-pull digger. Although alvarezsaurids were long tailed, however, a long tail is typical of diapsid reptiles, so this in itself was not necessarily a specific adaptation to digging in alvarezsaurids. That being said, alvarezsaurids did have a few unusual features of the tail that may shed light on their lifestyle, and Meso et al. (2021) provided a detailed description and functional interpretation of alvarezsaurid tail anatomy.
For one, alvarezsaurids were long-tailed even for theropods; relative to their body size, they probably had among the longest tails of any maniraptoran. For another, many features of their tail vertebrae suggest that their tails were more flexible from side to side than those of most other theropods. Probably to compensate for this increase in length and mobility, the muscles along the top of the tail that would have helped hold it aloft appear to have been well developed in alvarezsaurids. Also abnormal among theropods is the fact that alvarezsaurid tail vertebrae were procoelous: the back end of each vertebra fit into a socket in the front end of the vertebra behind it. (In typical theropods, the joint surfaces of the tail vertebrae tend to be flat or nearly so.) This may have been another way of reinforcing the tail, giving it further mobility without a correspondingly high risk of dislocating the vertebrae.
 |
| Reconstructed tail musculature of an early alvarezsaur (A), a patagonykine alvarezsaurid (B), and a parvicursorine alvarezsaurid (C), from Meso et al. (2021). |
Meso et al. note that overall configuration of the alvarezsaurid tail exhibits similarities to that of the extant aardvark (Orycteropus afer), which also has large muscles running along the top of a long (by mammal standards), strong, and flexible tail. In aardvarks, the tail probably serves an important role as a brace while digging (Endo et al., 2013). This could have also been the case in alvarezsaurids, but Meso et al. suggest that their characteristic tail morphology might have provided another adaptive benefit. As I discussed previously, one of the many strange features seen in alvarezsaurids is their apparent specialization towards running, and it is likely that they relied on this as a means to escape predation. Meso et al. posit that their increased tail flexibility gave alvarezsaurids a smaller turning radius, granting them a further advantage in avoiding pursuers.
 |
| Diagram showing how the flexible tail of alvarezsaurids might have helped them avoid predators (with a successful escape making use of a reduced turning radius shown in B), from Meso et al. (2021). |
Creatures of the Night
How an animal perceives the world naturally has a major influence on how it interacts with its environment and the other organisms in it, and Choiniere et al. (2021) offered some considerable insight into this aspect of alvarezsaurid biology. Their study was not focused solely on alvarezsaurs, but compared the proportions of the scleral ring (a bony ring embedded in the eyeball of many vertebrates, though not in mammals) and the anatomy of the inner ear in a wide variety of Mesozoic dinosaurs to those of extant birds and other reptiles. However, the results they found for the alvarezsaurid Shuvuuia (one of the few alvarezsaurs in which both of these elements are well preserved) were especially intriguing.
The anatomy of both the scleral ring and inner ear in Shuvuuia strongly suggests that it was primarily active at night. What's more, its inner ear exhibits an extreme morphology similar to that of the extant western barn owl (Tyto alba), a nocturnal predator that famously uses sensitive hearing to detect prey. It's important to emphasize here (as I've seen some popular retellings of this study misunderstand this) that the observed similarities between the inner ear of Shuvuuia and barn owls are due to the structure of the ear itself, not related to ear asymmetry.
Barn owls and their close relatives in the group Tytonidae are one lineage of owls that have evolved asymmetrical ears, which help them pinpoint the location of their prey. Ear asymmetry in owls can take several different forms, but in tytonids, the left ear opening is generally positioned higher than the right. However, this asymmetry is not reflected in their skull (Norberg, 2002). There are owls (such as those in the genus Aegolius) that do have asymmetrical skulls, but tytonids are not among them. Choiniere et al. also found no evidence of skull asymmetry in Shuvuuia. Might alvarezsaurids have instead had asymmetry in the soft tissue morphology of their ears, like barn owls? That seems like a possibility, but we may not be able to determine one way or another from their bony anatomy.
 |
| The back of the skull and inner ear of Shuvuuia (A–C) compared to that of a western barn owl (D–F), from Choiniere et al. (2021). |
As noted by Choiniere et al., a combination of a nocturnal lifestyle with specialized hearing is widespread in tetrapods, especially in mammals, so the fact that Shuvuuia exhibits evidence of these traits does not by itself indicate that alvarezsaurids were myrmecophagous (feeding mostly on social insects). However, there is indeed an extant myrmecophage that often forages at night primarily by acoustic cues, the bat-eared fox (Otocyon megalotis) (Renda and le Roux, 2017). Therefore, I consider these new findings on alvarezsaurid sensory biology very much compatible with the hypothesis that they were myrmecophages. (In fact, I was aware of Choiniere et al.'s research before its formal publication, and deliberately inserted a reference to the bat-eared fox into my last alvarezsaur post as oblique foreshadowing.)
 |
| A bat-eared fox, photographed by Yathin S Krishnappa, under CC BY-SA 3.0. This species feeds mainly on termites, which it locates using its sensitive hearing. Perhaps alvarezsaurids did the same? |
The Incredible Shrinking Alvarezsaurs
My previous post also mentioned small body size as as a curious evolutionary trend in later alvarezsaurids, and Qin et al. (2021) investigated this phenomenon in more detail. First of all, by inferring growth patterns based on the bone microstructure of individual alvarezsaur specimens, they confirmed that some alvarezsaurids, such as Albinykus and Xixianykus, had adult body masses of only about 1 kg or less. (It's worth noting though that one of the smallest known alvarezsaurids, the type specimen of Parvicursor, was subsequently reinterpreted as a juvenile instead of an adult by Averianov and Lopatin, 2021.)
Interestingly, Qin et al. indicated that Albinykus and Xixianykus had very different growth strategies from one another despite being of similar sizes. Whereas the type specimen of Albinykus had essentially stopped growing by the time it was three years old, the type specimen of Xixianykus grew more slowly but across a longer period of time, living for over a decade. This diversity in alvarezsaurid growth patterns was also observed more recently by D'Emic et al. (2023).
When Qin et al. plotted alvarezsaur body size across the evolutionary history of these dinosaurs, they found that not only were later alvarezsaurids smaller overall than their ancestors, but their small body size evolved very quickly in the early Late Cretaceous, about 90 million years ago. What may have caused this sudden miniaturization? Qin et al. point out that this event would have shortly followed the estimated diversification of both ants and termites during the Cretaceous, which may have created the opportunity for dedicated myrmecophagy to evolve in alvarezsaurs. Specialized insectivory as a driver of alvarezsaurid size reduction would also align with the median body size of insectivorous land vertebrates being consistently smaller than those with other dietary habits, a pattern that has been shown to hold true across different groups and biomes (Cooke et al., 2022).
 |
| Evolution of alvarezsaur body size over time, from Qin et al. (2021). |
When All You Have is a Pick...
As I hope I reviewed thoroughly in my last alvarezsaur post, there are many features of alvarezsaurid forelimbs that suggest they functioned in digging. However, just how effective their claws would have been at this task had not been quantitatively tested until recently. Qin et al. (2023) applied finite element analysis (FEA) to alvarezsaur hand claws to compare how they performed under different scenarios. (This study also looked at the claws of therizinosaurs, but this post is not about them.)
For each claw studied, three scenarios were tested: piercing (puncturing a substrate), pulling (using the underside of the claw to pull an object downward), and scratching (dragging the tip of the claw through a substrate), with the last being considered analogous to digging. When compared to a variety of mammals representing a range of claw morphologies and functions, the claws of alvarezsaurids were found to behave akin to those of pangolins: experiencing relatively low stress and similar stress distributions across all three scenarios. On the surface, it may sound surprising that such seemingly specialized claws would be well suited to multiple tasks in this way, but as Qin et al. point out, this makes a lot of sense if alvarezsaurids were diggers. Hook-and-pull digging in particular requires piercing, pulling, and scratching motions, so it would be potentially advantageous for animals adapted for this behavior to excel at all three.
Furthermore, Qin et al. found that the claws of alvarezsaurids performed better at scratching than those of earlier, non-alvarezsaurid alvarezsaurs, which had hands more broadly similar to those of typical theropods. This may suggest that increased digging specializations were specifically being selected for during alvarezsaur evolution.
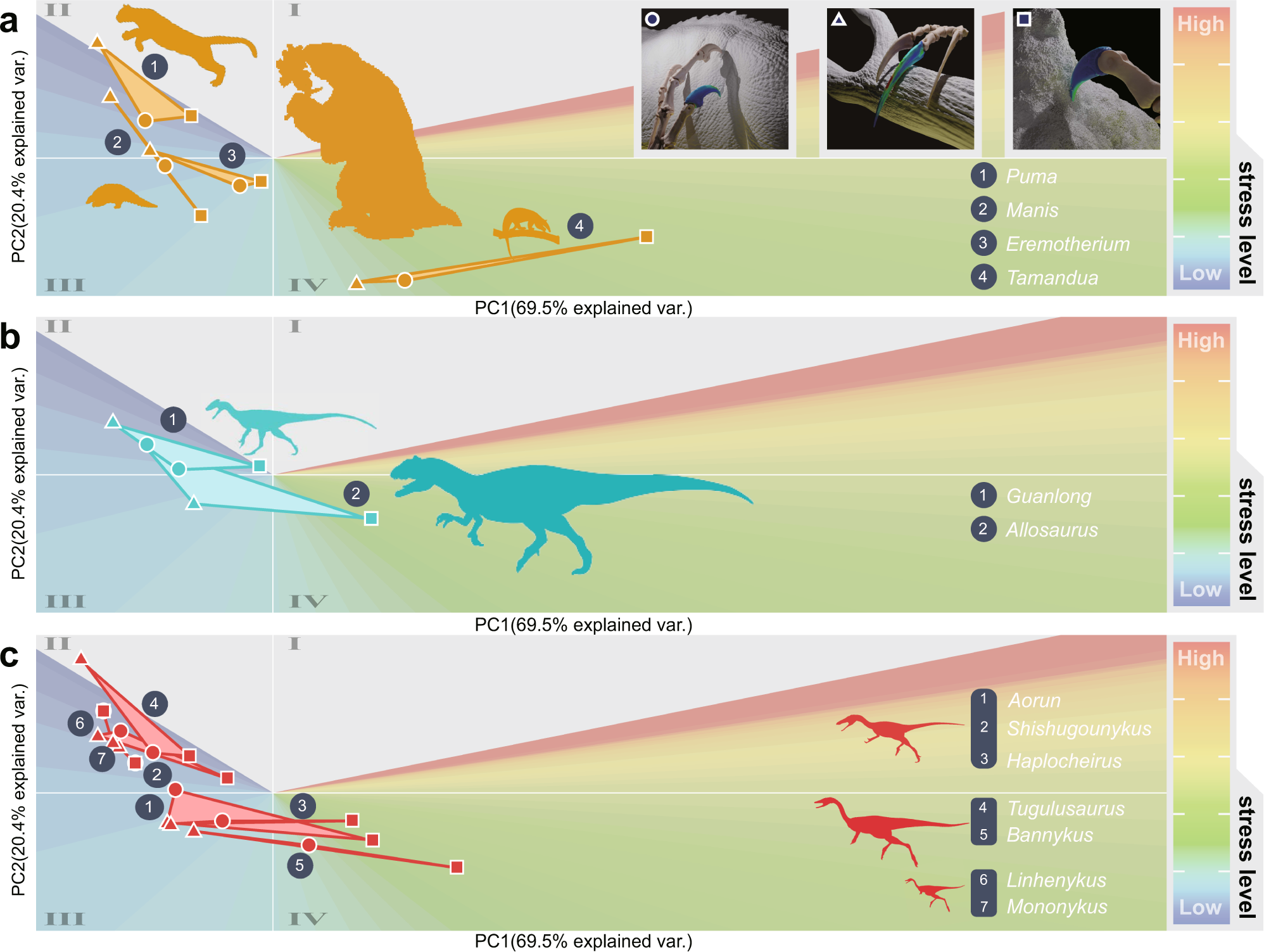 |
| Claw performance under different simulated scenarios in a selection of mammals (a), non-alvarezsaurian theropods (b), and alvarezsaurs (c), from Qin et al. (2023). What surprises me the most is that the fields for the pangolin and the tamandua plot so far apart, especially considering that, according to the supplementary material, the pangolin species studied was the tree pangolin (Phataginus ["Manis"] tricuspis), which, like tamanduas, spends a lot of time in trees. Maybe there is more functional variation among these superficially similar myrmecophages than commonly appreciated. |
It makes me happy to see so much new alvarezsaur research being done lately, and new advances have been made in understanding the digging abilities of extinct mammals as well (Nakai and Fujiwara, 2023), which could perhaps be applied to alvarezsaurs in the future. I'm certainly pleased that essentially all of these new alvarezsaur studies either reinforce or are consistent with the hypothesis that alvarezsaurids were myrmecophages. However, what excites me most of all is that I might have had a very small hand in bringing some of this science to fruition. I have it on good authority from Qin Zichuan that he and his colleagues' study on alvarezsaurid claw biomechanics was directly inspired by my previous alvarezsaur post, in which I idly suggested some potential directions for future investigations into alvarezsaurid ecology. For my humble blog post, having stimulated actual scientific research on these remarkable dinosaurs is just about the highest honor that I can conceive of.
 |
| Excerpt from the acknowledgements of Qin et al. (2023). |
References
- Agnolín, F.L., J.-C. Lu, M. Kundrát, and L. Xu. 2022. Alvarezsaurid osteology: new data on cranial anatomy. Historical Biology 34: 443–452. doi: 10.1080/08912963.2021.1929203
- Averianov, A.O. and A.V. Lopatin. 2021. A re-appraisal of Parvicursor remotus from the Late Cretaceous of Mongolia: implications for the phylogeny and taxonomy of alvarezsaurid theropod dinosaurs. Journal of Systematic Palaeontology 19: 1097–1128. doi: 10.1080/14772019.2021.2013965
- Averianov, A.O. and A.V. Lopatin. 2022a. The second taxon of alvarezsaurid theropod dinosaurs from the Late Cretaceous Khulsan locality in Gobi Desert, Mongolia. Historical Biology 34: 2125–2136. doi: 10.1080/08912963.2021.2000976
- Averianov, A.O. and A.V. Lopatin. 2022b. A new alvarezsaurid theropod dinosaur from the Upper Cretaceous of Gobi Desert, Mongolia. Cretaceous Research 135: 105168. doi: 10.1016/j.cretres.2022.105168
- Averianov, A.O. and H.-D. Sues. 2021. New material and diagnosis of a new taxon of alvarezsaurid (Dinosauria, Theropoda) from the Upper Cretaceous Bissekty Formation of Uzbekistan. Journal of Vertebrate Paleontology 41: e2036174. doi: 10.1080/02724634.2021.2036174
- Choiniere, J.N., J.M. Neenan, L. Schmitz, D.P. Ford, K.E.J. Chapelle, A.M. Balanoff, J.S. Sipla, J.A. Georgi, S.A. Walsh, M.A. Norell, X. Xu, J.M. Clark, and R.B.J. Benson. 2021. Evolution of vision and hearing modalities in theropod dinosaurs. Science 372: 610–613. doi: 10.1126/science.abe7941
- Cooke, R., W. Gearty, A.S.A. Chapman, J. Dunic, G.J. Edgar, J.S. Lefcheck, G. Rilov, C.R. McClain, R.D. Stuart-Smith, S.K. Lyons, and A.E. Bates. 2022. Anthropogenic disruptions to longstanding patterns of trophic-size structure in vertebrates. Nature Ecology and Evolution 6: 684–692. doi: 10.1038/s41559-022-01726-x
- D'Emic, M.D., P.M. O'Connor, R.S. Sombathy, I. Cerda, T.R. Pascucci, D. Varricchio, D. Pol, A. Dave, R.A. Coria, and K.A. Curry Rogers. 2023. Developmental strategies underlying gigantism and miniaturization in non-avialan theropod dinosaurs. Science 379: 811–814. doi: 10.1126/science.adc8714
- Endo, H., K. Mori, D. Koyabu, S. Kawada, T. Komiya, T. Itou, H. Koie, M. Kitagawa, and T. Sakai. 2013. Functional morphology of the aardvark tail. Anatomia, Histologia, Embryologia 42: 90–98. doi: 10.1111/j.1439-0264.2012.01169.x
- Fowler, D.W., J.P. Wilson, E.A. Freedman Fowler, C.R. Noto, D. Anduza, and J.R. Horner. 2020. Trierarchuncus prairiensis gen. et sp. nov., the last alvarezsaurid: Hell Creek Formation (uppermost Maastrichtian), Montana. Cretaceous Research 116: 104560. doi: 10.1016/j.cretres.2020.104560
- Freimuth, W.J. and J.P. Wilson. 2021. New manual unguals of Trierarchuncus prairiensis from the Hell Creek Formation, Montana, and the ontogenetic development of the functional alvarezsaurid hand claw. Cretaceous Research 119: 104698. doi: 10.1016/j.cretres.2020.104698
- Meso, J.G., Z. Qin, M. Pittman, J.I. Canale, L. Salgado, and V. Díez Díaz. 2021. Tail anatomy of the Alvarezsauria (Theropoda, Coelurosauria), and its functional and behavioural implications. Cretaceous Research 124: 104830. doi: 10.1016/j.cretres.2021.104830
- Nakai, D. and S. Fujiwara. 2023. Fossorial mammals emphasise the forelimb muscle moment arms used for digging: new indices for reconstruction of the digging ability and behaviours in extinct taxa. Journal of Anatomy 242: 846–861. doi: 10.1111/joa.13815
- Norberg, R.Å. 2002. Independent evolution of outer ear asymmetry among five owl lineages; morphology, function and selection. Pp. 329–342, in I. Newton, R. Kavanagh, J. Olsen, and I. Taylor (eds.), Ecology and Conservation of Owls. CSIRO Publishing, Clayton.
- Qin, Z., Q. Zhao, J.N. Choiniere, J.M. Clark, M.J. Benton, and X. Xu. 2021. Growth and miniaturization among alvarezsauroid dinosaurs. Current Biology 31: 3687–3693. doi: 10.1016/j.cub.2021.06.013
- Qin, Z., C.-C. Liao, M.J. Benton, and E.J. Rayfield. 2023. Functional space analyses reveal the function and evolution of the most bizarre theropod manual unguals. Communications Biology 6: 181. doi: 10.1038/s42003-023-04552-4
- Renda, S. and A. le Roux. 2017. The sensory ecology of prey detection in the bat-eared fox (Otocyon megalotis). Behaviour 154: 227–240. doi: 10.1163/1568539X-00003419














