
Greetings! I am Skull, a fossil cast of the skull of the deinonychosaurian maniraptor Tsaagan mangas. That happens to be the only part of Tsaagan mangas known to science aside from a few connected neck vertebrae. But I digress. Albertonykus has enlisted my help in providing a thorough and hopefully comprehensive intro of the group of dinosaurs known as the maniraptors to which this blog slash web comic is dedicated.
What is a maniraptor? As the header of the blog should imply, a maniraptor is any member of the most inclusive clade containing Passer domesticus but not Ornithomimus edmontonicus, but that may be somewhat difficult to understand for some. What it means is that, to qualify as a maniraptor, a creature must share a common ancestor (which would itself be a maniraptor) with a house sparrow that is more recent than the one they share with the Ornithomimus edmonticus. (The house sparrow itself, of course, is also a maniraptor, because it's "closer" to itself than to Ornithomimus edmonticus. That sounds paradoxical, but never mind; it should be understandable enough.)
That's all very well for the definition of "maniraptor", but how do you recognize a maniraptor when you see one? I direct you to the cladogram below (by Dr. Thomas Holtz; used with permission). Find "Maniraptora" circled in blue.
We maniraptors share with each other enlarged forelimbs, large bony sternums (breast bones), and well-developed semilunate carpals (a special bone in the wrist that allows maniraptors to tuck their forelimbs up against our bodies). We might also count backwards-pointing pubes (that's the plural of pubis, which is a hip bone), but, as you'll see in a moment, that story is complicated. Many maniraptors have backwards-pointing pubes, but it's not certain if we all inherited them from our original maniraptor ancestor. Either way, our long folding arms are useful for catching prey and also for keeping our feathers off the ground, preventing them from being soiled. You see, all us maniraptors have feathers, or at least protofeather fuzz, which is a trait we share with the rest of the Coelurosauria (a group that includes compsognathids, tyrannosauroids, ornithomimosaurs, and us; see the cladogram). Another thing special about us is that nearly all of us aren't hunters of big dinosaurs. Many of us eat plants, invertebrates, small vertebrates, or all three. Perhaps I shouldn't have said "us", because the main exception to this among the maniraptors is... well, I'll get to that later.
Enough is known about most maniraptors to split us into several groups, which I'll introduce one by one.
Therizinosaurs
 |
| From shortest to longest: Beipiaosaurus inexpecticus, Falcarius utahensis, and Therizinosaurus cheloniformis. |
Ebeff the Therizinosaurus cheloniformis is one of these. Therizinosaurs are really freaky-looking dinosaurs, what with their tiny heads on their long necks and gigantic claws on their hands! They are definitely plant eaters, because they are most likely too ponderous and slow to catch other animals, although their large claws would have made nice defensive weapons. Most therizinosaurs we know have backwards-pointing pubes and probably cheeks, both of which are features of other (unrelated) plant-eating dinosaurs. Both of these characteristics help therizinosaurs get the most out of their vegetarian diet: backwards-pointing pubes give them more room for bigger and longer guts (which house micro organisms for digesting plant matter), and cheeks help them retain food in the mouth so it doesn't slop everywhere and go to waste! However, the most basal therizinosaur known from complete remains, Falcarius utahensis, has a forward-pointing pubis and seems to be a decent runner, so it is probably more omnivorous than its derived relatives. As far as their plumage is concerned, therizinosaurs don't have the modern-style true feathers of today's birds. They have long shaggy protofeathers as well as a type of feather probably unique to them called EBFFs, or Elongated Broad Filamentous Feathers. EBFFs resemble porcupine quills and may have a display function. Most known therizinosaurs come from the Cretaceous, but phylogenetic bracketing tells us their origins lie back in the Jurassic. Indeed, an ambiguous lower jaw bone, given the name Eshanosaurus deguchiianus, appears to be that of a really old therizinosaur. Therizinosaurs are sheep- to Tyrannosaurus-rex-sized, making Therizinosaurus cheloniformis the largest known maniraptor.
Alvarezsauroids
 |
| From shortest to longest: Parvicursor remotus, Albertonykus borealis, and Haplocheirus sollers. |
This group has always been fairly strange. For one thing, no one really had any idea what they were! The first specimens were thought to be flightless avialians (more on those later), and later it was suggested that they may be more basal maniraptors or related to the ornithomimosaurs (ostrich dinos). But at the beginning of this year the basal alvarezsauroid Haplocheirus sollers was found (back in the Jurassic!), and we now know a good deal more about the history of this group. For a start, they do appear to maniraptors. They started out looking like typical generalist maniraptors with three-fingered hands, long arms, small teeth suitable to an omnivorous diet, and long legs. However, derived alvarezsauroids, the alvarezsaurids, have very short arms and are smaller in body size than their ancestors. Their third and second fingers are tiny and clawless, but they retain a big thumb claw. In spite of their length, those arms are very powerful, and it's been speculated that alvarezsaurids use them to pound into rotting wood and insect nests in search of food. Alvarezsaurids (especially the members of the alvarezsaurid group Parvicursorinae) are very good runners with their long, thin lower legs, and they have weakly backward-pointing pubes. So far, we don't really know what alvarezsauroid plumage was like, because there aren't any good fossils of alvarezsauroids that preserve integument. (To be fair, there's a specimen of Shuvuuia deserti that comes with very badly preserved protofeathers - can't really blame it though, it happened to live in a place that didn't preserve soft tissues well, in contrast to the creatures that wound up on lakebeds in the Yixian.) Alvarezsauroids are typically quite small. Even the larger basal ones are no more than three meters long.
Oviraptorosaurs
 |
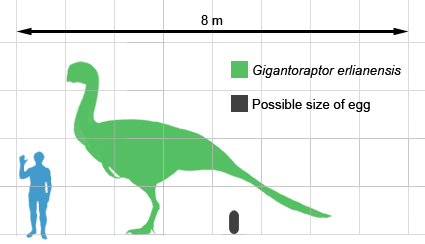
| From shortest to longest: Caudipteryx zoui, Citipati osmolskae, and Gigantoraptor erlianensis. |
Remex the Caudipteryx zoui is one of these. Many of you probably know how the oviraptorosaur Oviraptor philoceratops was discovered on a nest of eggs, earning it a name that means "egg thief, fond of horned dinosaurs", but was later found to have been brooding its own nest. We don't know if earlier maniraptors like alvarezsauroids and therizinosaurs brood, but we know that oviraptorosaurs definitely do. Also, oviraptorosaurs definitely have true feathers. They have their true feathers on the wings and tail. However, oviraptorosaurs can't fly, so what are their feathers for? They can help oviraptorosaurs cover their nests better in the aforementioned brooding behavior. Oviraptorosaurs may also use their feathers for showing off. Caudipteryx zoui, for example, waves little hand flags around - its wing feathers are concentrated on the second finger, while there aren't any true feathers on its arms! Feathers probably aid small oviraptorosaurs in a behavior called WAIR, short for Wing-Assisted Incline Running. By flapping their wings back and forth, small oviraptorosaurs can run up the sides of trees and escape from predators. Basal oviraptorosaurs have a few small teeth, and they eat plants or insects. Derived oviraptorosaurs, however, have no teeth at all, just keratinous beaks. They are most likely omnivores or predators of small vertebrates, as lizards have been found as their stomach contents and remains of smaller maniraptors have been found near their nests, possibly signifying the leftovers of eaten prey. (And, of course, just because they don't cannibalize their own eggs doesn't mean oviraptorosaurs can't eat the eggs of other animals.) Oviraptorosaurs have forward-pointing pubes, and many have adaptations to fast running such as long legs and pinched feet. Oviraptorosaurs range from turkey-sized to as large as the tyrannosauroid Albertosaurus sarcophagus.
Henceforth, all other maniraptors belong to the group Eumaniraptora. Eumaniraptors include deinonychosaurs and avialians, and all the characters (well, almost all) in this web comic were eumaniraptors before Remex and Ebeff showed up. This calls for a more detailed cladogram (again, by Dr. Thomas Holtz; also used with permission).
Us eumaniraptors have a first toe placed low on the foot and a retractable second toe we hold off the ground usually when walking. This keeps the second claw nice and sharp, which might be good for climbing (up trees, but some of us began climbing other things later on). (Modern birds don't have retractable toes anymore and instead grab things with a reversed first toe.) We also tend to have backwards-pointing pubes, which might be a result of our knee-driven running. Like oviraptorosaurs, we eumaniraptors have true feathers on the wings and tail, and we use them for similar purposes. However, we also have true feathers on the legs, too (ancestrally, anyways; some of us, again including modern birds, have lost that feature). Furthermore, we can actually fly! It's been suggested that those of us who can't (such as me... well, the species I was modeled after, really) are what is called secondarily flightless, which means we are flightless animals with flighted ancestors with flightless ancestors. The same thing is true of flightless modern birds and many well-known deinonychosaurs... which is the cue for another intro.
Deinonychosaurs
 |
| From shortest to longest: Microraptor zhaoianus, Troodon formosus, and Deinonychus antirrhopus. |
I'm a deinonychosaur, and so are Ostrom the Deinonychus antirrhopus and Zahavi the Jinfengopteryx elegans. The public knows our group quite well as "raptors", especially the members of Dromaeosauridae, which include both me and Ostrom. (Zahavi belongs to the Troodontidae, but you can worry about that later.) The public is more familiar with "typical" dromaeosaurids such as Deinonychus antirrhopus or me (or more likely my close relative Velociraptor mongoliensis, really), but the thing is we are far from being typical eumaniraptors! We can't fly, for example. Also, we attack and eat prey comparable in size to or larger than ourselves, which is something (most) other maniraptors don't do! (It's been suggested that at least some of us did this in groups, no less.) That means we may not be just secondarily flightless, but secondarily carnivorous as well! (As maniraptors belong to the coelurosaurs. Coelurosaurs were ancestrally carnivorous, as evidenced by meat-eating tyrannosauroids and compsognathids.) Some of us "typical" dromaeosaurids have grown so big (like Utahraptor ostrommaysorum) that they can no longer climb trees and use their retractable claws to climb up big plant eaters instead! Other dromaeosaurids can generally still fly and are much smaller than we are, but one group, the unenlagiines, largely returned to the ground after reaching (possibly flying) to the southern hemisphere and diversifying. Unenlagiines have long narrow snouts, so they probably eat fish and other small animals. At least one unenlagiine, Austroraptor cabazai, is nearly as large as Utahraptor ostrommaysorum.
The other main branch of the Deinonychosauria, the Troodontidae, are generally smaller (pigeon- to wolf-size) and faster than us. (Yes, sadly, in spite of media portrayals and the genus name of Velociraptor mongoliensis meaning "fast thief", we "typical" dromaeosaurids are a lot less built for running than troodonts, oviraptorosaurs, alvarezsauroids, or even our own ancestors, the basal dromaeosaurids!) Troodonts might have abandoned flight early on (even the most basal known troodont, Anchiornis huxleyi, shows signs of losing aerial capacity) and they usually have rather short arms. Their teeth are small and serrationless, so their dietry habits are more like that of other maniraptors: plants and small-bodied animals. (Jinfengopteryx elegans, for example, preserves seeds in its stomach.) Finally, the most derived troodonts have pubes that point forward rather than backwards like the rest of us. (That makes their pubes secondarily forward pointing!)
Avialians
 |
| From shortest to longest: Passer domesticus, Archaeopteryx lithographica, and Hesperornis regalis. |
Savape the Archaeopteryx lithographica and Dinky the house sparrow are both avialians. And so we come to the last major group of maniraptors. When people say birds, they usually refer to this maniraptor group. All modern birds belong to a group of avialians called the neornithines, but there are many kinds of Mesozoic avialians, many of them quite different from modern ones. The most basal avialians known are the tiny Jurassic scansoriopterygids, which are extremely weird with their elongated third fingers and ribbon-shaped tail feathers. We don't know much about the scansoriopterygids, but we know a lot about a group of Cretaceous birds called confuciusornithids, because there are probably over a thousand confuciusornithid fossils out there. Like modern birds, confuciusornithids have the tips of their tails fused into a bony feather-anchoring base called the pygostyle (which has also been independently evolved by some oviraptorosaurs and the therizinosaur Beipiaosaurus inexpectus). Unlike modern birds, confuciusornithids don't have a feather fan on their tails, but a pair of very long feathers. (Some confuciusornithid specimens do not have the long feathers; this may represent gender or age differences, or simply moulting individuals.) Confuciusornithids have toothless beaks which they use to catch fish as they fly over lakes, but they evolved these separately from modern birds. The enantiornithines were probably the most successful avialian group in the Mesozoic, spanning a wide variety of sizes (from sparrow- to vulture-sized) and habits (fish eaters, birds of prey, seed eaters, insect eaters, and others). Most enantiornithines don't have beaks or tail fans (but there are exceptions in both cases) and most retain teeth, protruding wing claws, and leg feathers, but they do have an alula, a feather on the thumb (also present in modern birds) that helps in steering and a fully backwards-pointing first toe. (Which isn't turned backwards in the most basal avialians and not fully reversed in confuciusornithids.) Most enantiornithines had two long tail feathers like confuciusornithids, but not to the same degree. Hesperornithines are diving birds, most of which have lost their forelimbs and are flightless. Although they have teeth, they do have tail fans and are closer to neornithines than any of the other groups mentioned so far. Hesperornithines swam with lobed feet like modern grebes instead of webbed ones. Neornithines also seem to have started out as waterfowl. When the K/Pg extinction struck, the neornithines alone among all dinosaurs survived the ensuing disaster, spawning the myriad forms we see now.
That's it for this presentation! I hope you had fun reading that, whether or not it was new to you. Next time, we return to comics, this time starring Remex and Ebeff!










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
It's "maniraptoran," not "maniraptor."
ReplyDeleteI'm aware that maniraptoran is more commonly used, but I've seen maniraptor as a more informal term, similar to how oviraptorosaur is just shortened to oviraptor on occasion. In any case, I only using it as a colloquial name for Maniraptora, not scientific terminology.
ReplyDeleteTherizinosaur claws were most likely not good for combat, too long.
ReplyDeleteI wouldn't rule out a defensive function entirely. Those claws are larger than is normal for theropods, but to be honest they don't look to me all that "crazy long": http://azhdarcho.com/Art/Paleoart/beipiaosaurus.htm
ReplyDeleteAnd I'm sure their use in defense is said to be possible by at least some paleontologists: http://www.geol.umd.edu/~tholtz/G104/lectures/104coelur.html
Even if defense wasn't the main fuction, that doesn't mean they couldn't still use them that way. Most of them being rather slow-moving herbivores a la ground sloths (which also have large claws), it may have been important for them to have some defense.
Will you update the opening sentence for Avialae?
ReplyDeleteSpinosegnosaurus77/SpongeBobFossilPants
I might, though editing old blog posts is not a priority of mine.
DeleteNormally I'd agree, but I don't consider this an "old post" so much as an intro for newcomers to the blog.
DeleteSpinosegnosaurus77/SpongeBobFossilPants
Eh, it's more an intro to what maniraptors are rather than to the blog itself, and as I note on my about page, on that front Wikipedia is a better alternative if you want a frequently updated source.
Deletei know my question isnt directly related to the topic here but i just wanna know your answer. How do we know for sure whether a trait evolved thru natural selection or thru genetic drift.
ReplyDelete